Metabolism
on-line - the virtual tutorial room
copyright © 2008 - 2015 David A Bender
Two very hyperketotic children
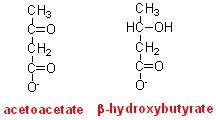 In a series
of experiments using isolated rat cardiomyocytes (heart muscle cells), the consumption
of oxygen was measured when various substrates were added. The figures below
show the consumption of oxygen when the cells were provided with 10 µmol
/L acetoacetate or beta-hydroxybutyrate as substrates (mean ± sd for
5 x replicate incubations).
In a series
of experiments using isolated rat cardiomyocytes (heart muscle cells), the consumption
of oxygen was measured when various substrates were added. The figures below
show the consumption of oxygen when the cells were provided with 10 µmol
/L acetoacetate or beta-hydroxybutyrate as substrates (mean ± sd for
5 x replicate incubations).
| substrate | nmol oxygen consumed /min /mg protein |
| acetoacetate | 240 ± 4 |
| beta-hydroxybutyrate | 270 ± 3 |
What conclusions can you draw from these results?
 If more
oxygen is consumed in the metabolism of beta-hydroxybutyrate than of acetoacetate,
this suggests that it is likely that the first step in metabolism of beta-hydroxybutyrate
involves its oxidation, presumably to acetoacetate, the reverse of the reaction
in the liver, in which acetoacetate is reduced to beta-hydroxybutyrate:
If more
oxygen is consumed in the metabolism of beta-hydroxybutyrate than of acetoacetate,
this suggests that it is likely that the first step in metabolism of beta-hydroxybutyrate
involves its oxidation, presumably to acetoacetate, the reverse of the reaction
in the liver, in which acetoacetate is reduced to beta-hydroxybutyrate:
We have already seen (in the exercise on Two boys with profound fasting hypoglycaemia and no ketone bodies) that the advantage of reducing acetoacetate to beta-hydroxybutyrate in the liver is that acetoacetate is unstable, and undergoes non-enzymic decarboxylation to acetone, which is metabolically more or less useless. Reduction to beta-hydroxybutyrate thus prevents loss of metabolic fuel in fasting and starvation.
What do you think is the advantage to extra-hepatic tissues of receiving beta-hydroxybutyrate rather than acetoacetate?
The oxidation of beta-hydroxybutyrate to acetoacetate involves reduction of NAD to NADH - this will provide an additional ~2.5 ATP per mol of beta-hydroxybutyrate metabolised compared with acetoacetate.
Reduction of acetoacetate to beta-hydroxybutyrate is thus a way of exporting reducing equivalents, and indirectly ATP, from the liver to extra-hepatic tissues.
SA is a one year old boy. Intermittently he has suffered metabolic crises, usually associated with mild infections and loss of appetite, that have resulted in hospitalisation because he became unconscious and was hyperventilating. On the most recent such hospital admission blood tests revealed the following results (data for control subjects of the same age show the 10th - 90th centile range and mean in brackets for 12 children fasted for 20 hours):
SA |
control children |
||
10th - 90th centile range |
mean |
||
| pH | 6.88 |
7.35 - 7.45 |
7.4 |
| pCO2 (mmHg) | 24 |
35 - 46 |
40.5 |
| PO2 (mmHg) | 90 |
85 - 105 |
95 |
| bicarbonate (mmol /L) | 4.0 |
21.0 - 29.0 |
25.0 |
| glucose (mmol /L) | 3.0 |
3.5 - 4.6 |
3.9 |
| lactate (mmol /L) | 1.3 |
0.85 - 1.8 |
1.3 |
| non-esterified fatty acids (mmol /L) | 3.0 |
0.6 - 1.3 |
0.9 |
| total ketone bodies (mmol /L) | 10.3 |
0.6 - 3.2 |
1.6 |
| beta-hydroxybutyrate (mmol /L) | 6.0 |
0.5 - 2.3 |
1.1 |
| non-esterified fatty acids : ketone bodies (ratio) | 0.3 |
0.3 - 1.4 |
0.8 |
| non-esterified fatty acids : beta-hydroxybutyrate (ratio) | 0.5 |
0.5 - 1.9 |
1.1 |
| beta-hydroxybutyrate : acetoacetate (ratio) | 1.4 |
1.9 - 3.1 |
2.5 |
| insulin (µU /mL) | 3.7 |
3.5 - 5.5 |
4.5 |
| glucagon (pg /mL) | 195 |
150 - 190 |
170 |
(From data reported by Bonnefont et al. Eur J Pediatr 150: 80-5, 1990, and Niezen-Koning et al. Eur J Pediatr 156: 870-3, 1997)
What conclusions can you draw from these results?
He is clearly acidotic, with low plasma bicarbonate and pCO2, but a normal pO2. This suggests a metabolic acidosis rather than respiratory acidosis, in which case pCO2 and bicarbonate would be expected to be higher than normal.
The acidosis seems be due to very high concentrations of ketone bodies - his plasma lactate (another common cause of metabolic acidosis) is within the normal range.
He is slightly hypoglycaemic, but his plasma concentrations of insulin and glucagon are appropriate for his plasma glucose concentration, so there is nothing to suggest an abnormality of secretion of, or responsiveness to, either insulin or glucagon.
His plasma non-esterified fatty acids and ketone bodies are both massively elevated; the ratios of non-esterified fatty acids : total ketone bodies and beta-hydroxybutyrate are at the lower end of the normal range. There results suggest that he is perfectly able to mobilise non-esterified fatty acids from adipose tissue triacylglycerol, and to synthesise synthesise ketone bodies from non-esterified fatty acids. It is most likely that his problem is in the utilisation of ketone bodies in extra-hepatic tissues.
The ratio of beta-hydroxybutyrate : acetoacetate is lower than normal. This might suggest impaired activity of beta-hydroxybutyrate dehydrogenase in the liver. However, it is equally likely that he has normal activity of beta-hydroxybutyrate dehydrogenase in both liver and peripheral tissues, but is unable to metabolise the resultant acetoacetate.
In the following experiments cultured skin fibroblasts were incubated with 10mmol /L [14C] beta-hydroxybutyrate or acetoacetate and the radioactivity in carbon dioxide was measured. Figures show nmol of carbon dioxide produced /min /mg protein, with mean ± sd for cultured fibroblasts from 10 control subjects being investigated for unrelated conditions. At the end of the incubation beta-hydroxybutyrate and acetoacetate were measured in the incubation medium.
substrate |
[14C]CO2 (dpm)
|
final beta-hydroxybutyrate
( mmol /L) |
final acetoacetate (mmol
/L)
|
|||
SA |
controls |
SA |
controls |
SA |
controls |
|
beta-hydroxybutyrate (10 µmol /L) |
27 |
1987 ± 125 |
0.1 |
4.5 ± 0.6 |
9.8 |
0.1 ± 0.05 |
acetoacetate (10 µmol /L) |
23 |
2240 ± 153 |
nd |
nd |
9.9 |
5.1 ± 0.8 |
nd = not detectable