Metabolism
on-line - the virtual tutorial room
copyright © 2008 - 2015 David A Bender
Experiments on glucose metabolism
So far, from various studies we have seen that (anaerobic) glucose metabolism leads to the production of 2 mol of lactate and 2 mol of ATP per mol of glucose consumed; the process also requires ATP and inorganic phosphate. The amount of glucose consumed, and of lactate formed, depends on the amount of ADP provided.
For the following studies, chicken breast muscle was homogenised in phosphate buffer at pH 7.4, then subjected to high speed centrifugation to remove cell debris, nuclei and mitochondria. The supernatant fraction (essentially cytosol, but also containing ribosomes and microsomes, was then dialysed against four changes of buffer to remove any endogenous substrates and other low molecular weight compounds. Each incubation in the following sets of experiments contained the dialysed homogenate from 1 g of fresh muscle tissue.
Experiments were conducted incubating this preparation with one of:
[14C-U]glucose at a specific activity of 1 µCi /mmol
[32P]ATP at a specific activity of 1 µCi /mmol
[32P]inorganic phosphate at a specific activity of 1 µCi /mmol
In the first set of experiments, incubations contained:
200 µmol glucose
200 µmol ATP
200 µmol ADP
200 µmol sodium phosphate
After incubation at 30C for 30 min, incubations were stopped by adding trichloroacetic acid to denature proteins, centrifuged and the supernatant was neutralised with sodium hydroxide. Aliquots of the supernatant were then subjected to high pressure liquid chromatography to measure both the amounts of various metabolites present and and the radioactivity in each. The results are shown as total amount of each metabolite and the specific radioactivity (µCi / mmol) - mean values ± sd from 3 x replicate incubations.
The following results were obtained:
metabolite |
amount |
specific radioactivity
(µCi /mmol) |
|||
µmol |
from [14C-U]glucose |
from [32P]ATP |
from [32P] inorganic phosphate |
||
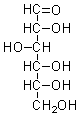 |
glucose |
100 ± 5 |
0.99 ± 0.02 |
- |
- |
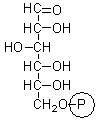 |
glucose 6-phosphate |
3.2 ± 0.4 |
0.98 ± 0.03 |
0.99 ± 0.03 |
0 |
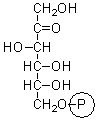 |
fructose 6-phosphate |
2.5 ± 0.3 |
0.99 ± 0.03 |
0.98 ± 0.03 |
0 |
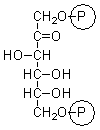 |
fructose 1,6-bisphosphate |
3.3 ±
0.7 |
0.99 ± 0.02 |
1.99 ± 0.04 |
0 |
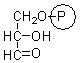 |
glyceraldehyde 3-phosphate |
94 ± 1.2 |
0.48 ± 0.03 |
0.99 ± 0.02 |
0 |
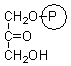 |
dihydroxyacetone phosphate |
98 ± 1.4 |
0.49 ± 0.02 |
1 |
0 |
ATP |
0 |
- |
- |
- |
|
ADP |
248 ± 5 |
- |
0 |
0 |
|
There was no detectable formation of lactate.
What conclusions can you draw from these results?
It is difficult to explain at this stage why there was no formation of lactate or ATP. Indeed, all of the ATP seems to have been converted to ADP, whereas in previous studies we have seen phosphorylation of ADP to ATP associated with glucose metabolism.
It is obvious from these results that glucose has been phosphorylated to glucose 6-phosphate; it then seems likely that glucose 6-phosphate has been isomerised to fructose 6-phosphate. Fructose 6-phosphate has then been phosphorylated to fructose 1,6-bisphosphate, and it looks as is fructose 1,6-bisphosphate has been cleaved to yield glyceraldehyde 3-phosphate and dihydroxyacetone phosphate. Neither of these three carbon compounds has undergone any further metabolism.
A further experiment was set up in which the amount of ATP per incubation was varied, and only glucose disappearance and the appearance of glyceraldehyde 3-phosphate and dihydroxyacetone phosphate were measured.
The results were as follows:
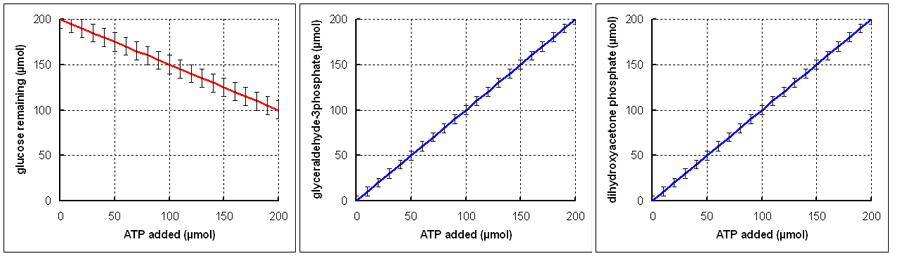
What conclusions can you draw from these results?
These graphs show that there is a requirement for 2 mol of ATP for each mol of glucose metabolised, and that 1 mol each of glyceraldehyde 3-phpsphate and dihydroxyacetone phosphate is formed per mol of glucose. These results thus confirm the conclusions drawn from the first experiment with dialysed muscle supernatant.
Like red blood cells, muscle preparations that are incubated anaerobically normally produce lactate from glucose.
Why do you think it may be that the dialysed muscle preparation did not form lactate?
The fact that the dialysed muscle preparation did not produce any lactate, although muscle incubated under anaerobic conditions forms lactate from glucose, suggests that there may be a low molecular weight compound that is required for the full pathway of glucose metabolism, and this has been lost in dialysis. A classical way of testing this hypothesis is to boil a sample of the tissue preparation before dialysis, centrifuge it to remove denatured protein, then add some of the supernatant to the incubation mixture. This preparation is commonly known as a kochsaft (from the German, meaning literally cooking or cooked juice).
A series of incubations was set up containing both the dialysed muscle preparation and either buffer or the kochsaft from boiled (but undialysed) muscle preparation, together with:
200 µmol glucose
200 µmol ATP
200 µmol ADP
200 µmol sodium phosphate
After incubation at 30C for 30 min, incubations were stopped by adding trichloroacetic acid to denature proteins, centrifuged and the supernatant was neutralised with sodium hydroxide. Aliquots of the supernatant were then subjected to high pressure liquid chromatography to measure glucose, lactate, ATP and ADP. The following results were obtained:
| control (+ buffer) | + kochsaft | |
| glucose | 100 |
100 |
| lactate | 0 |
80 |
| ATP | 0 |
400 |
| ADP | 300 |
0 |
What conclusions can you draw from these results?
The control incubations show the same results as before - no lactate is formed and 2 mol of ATP are consumed per mol of glucose metabolised. However, with the kochsaft added there is now formation of lactate, and instead of net consumption of ATP there is phosphorylation of ADP to ATP.
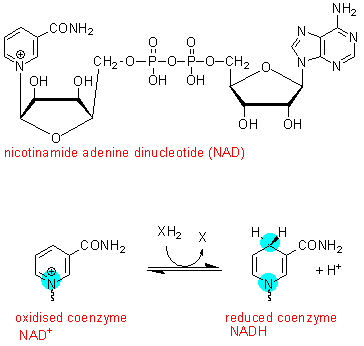 The obvious
next step is to discover what it is in the kochsaft that is required for the
complete metabolism of glucose to lactate. After a number of experiments it
was discovered that the compound concerned is nicotinamide adenine dinucleotide
(NAD), a coenzyme that undergoes oxidation and reduction reactions.
The obvious
next step is to discover what it is in the kochsaft that is required for the
complete metabolism of glucose to lactate. After a number of experiments it
was discovered that the compound concerned is nicotinamide adenine dinucleotide
(NAD), a coenzyme that undergoes oxidation and reduction reactions.
Using uv absorption spectrometry it is easy to follow redox reactions using NAD or NADH - both absorb uv light at 259 nm, but the reduced coenzyme (NADH) also absorbs at 340 nm. This means that an oxidation reaction can be followed by measuring the increase in absorbance at 340 nm, and a reduction reaction by measuring the disappearance of absorbance at 340 nm.
A series of incubations was set up containing the dialysed muscle preparation and varying amounts of NAD, together with:
200 µmol glucose
200 µmol ATP
200 µmol ADP
200 µmol sodium phosphate
 After
incubation at 30C for 30 min, incubations were stopped by adding trichloroacetic
acid to denature proteins, centrifuged and the supernatant was neutralised with
sodium hydroxide. Aliquots of the supernatant were then subjected to high pressure
liquid chromatography to measure glucose, lactate, NAD and NADH. An initially
unidentified compound was also found in the incubation mixtures, which was later
identified as pyruvate.
After
incubation at 30C for 30 min, incubations were stopped by adding trichloroacetic
acid to denature proteins, centrifuged and the supernatant was neutralised with
sodium hydroxide. Aliquots of the supernatant were then subjected to high pressure
liquid chromatography to measure glucose, lactate, NAD and NADH. An initially
unidentified compound was also found in the incubation mixtures, which was later
identified as pyruvate.
The following results were obtained:
What conclusions can you draw from these results?
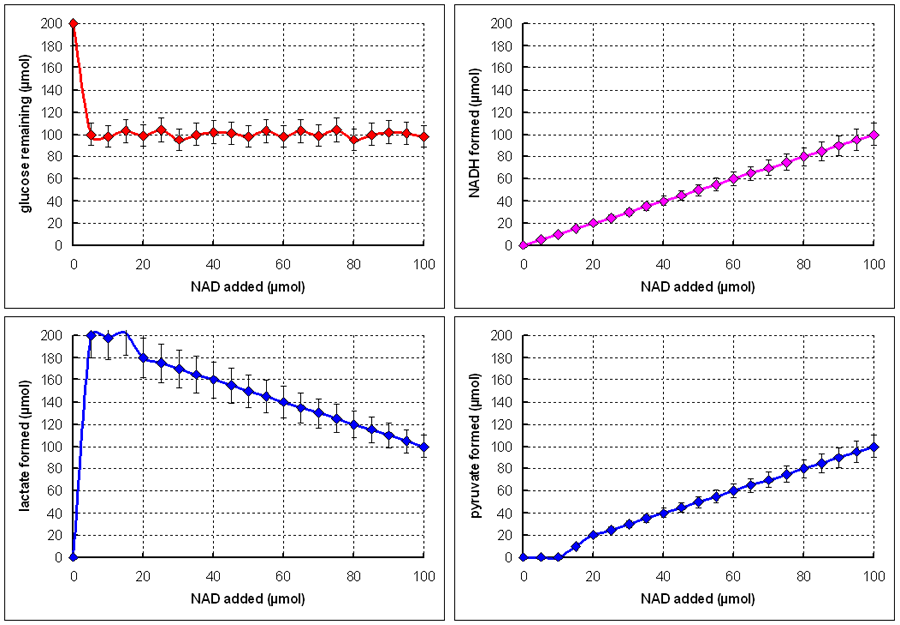
Even a very small amount of NAD seems to be adequate to permit full consumption of glucose (limited presumably by the amount of either ADP or ATP added). This suggests that NAD has a catalytic role on the pathway of glucose metabolism. However, at the end of the incubations almost al of the NAD had been reduced to NADH.
When
only a small amount of NAD was added, all of the glucose consumed was converted
to lactate (2 mol of lactate per mol of glucose consumed). However, as more
NAD was added, an increasing proportion of the glucose was metabolised to pyruvate
rather than lactate. Note the the sum of lactate + pyruvate formed is more or
less constant, at 2 mol of (lactate or pyruvate) per mol of glucose consumed.
What is the relationship between lactate and pyruvate, and how might they be interconverted?
You can either say that lactate is the product of reduction of pyruvate, or that pyruvate is the product of oxidation of lactate - they form a redox pair.
What would you expect to observe if you incubated the dialysed muscle supernatant preparation with lactate and NAD?
What would you expect to observe if you incubated the dialysed muscle supernatant preparation with pyruvate and NADH?
If you incubated with lactate and NAD, you would expect to see stoichiometric loss of lactate, appearance of pyruvate and appearance of NADH (detectable by its absorbance at 240 nm)
If you incubated with pyruvate and NADH, you would expect to see stoichiometric appearance of lactate, disappearance of NADH and disappearance of pyruvate.
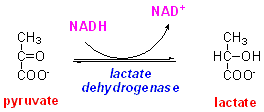 There is an
enzyme, lactate dehydrogenase, that catalyses the interconversion of lactate
and pyruvate, linked to utilisation of NAD and NADH:
There is an
enzyme, lactate dehydrogenase, that catalyses the interconversion of lactate
and pyruvate, linked to utilisation of NAD and NADH:
This suggests that NAD is reduced to NADH in the metabolism of glucose to form pyruvate, then lactate is formed to re-oxidise the NADH back to NAD.
The same set of incubations was performed using only 20 µmol of ATP, rather than 200 µmol. The results were the same.
What conclusion can you draw from this?
We already know that when glucose is metabolised to lactate there is a net yield of 2 ATP per glucose metabolised. EWe also know that there is a cost of 2 x ATP to convert glucose to fructose 1,2-bisphosphate. Obviously, since only a small amount of ATP (20 µmol in this experiment) is required to permit the metabolism of much more glucose (100 µmol in this experiment), then the total amount of ATP formed in the metabolism of 1 mol of glucose must be more than 2 mol of ATP formed. Not only the ADP added initially but also that resulting from phosphorylation of glucose to glucose 6-phosphate and fructose 6-phosphate to fructose 1,6-bisphosphate, is also phosphorylated to ATP.
This suggests that altogether there is probably formation of 4 mol of ATP per mol of glucose metabolised to lactate.
We can now return to our experiments with radioactive glucose, ATP and inorganic phosphate.
Experiments were conducted incubating an amount of the dialysed muscle supernatant preparation equivalent to 1 g of fresh muscle tissue with one of:
[14C-U]glucose at a specific activity of 1 µCi /mmol
[32P]ATP at a specific activity of 1 µCi /mmol
[32P]inorganic phosphate at a specific activity of 1 µCi /mmol
In the first set of experiments, incubations contained:
200 µmol glucose
20 µmol ATP
20 µmol NAD
200 µmol ADP
200 µmol sodium phosphate
After incubation at 30C for 30 min, incubations were stopped by adding trichloroacetic acid to denature proteins, centrifuged and the supernatant was neutralised with sodium hydroxide. Aliquots of the supernatant were then subjected to high pressure liquid chromatography to measure both the amounts of various metabolites present and and the radioactivity in each. The results are shown as total amount of each metabolite and the specific radioactivity (µCi / mmol) - mean values ± sd from 3 x replicate incubations.
The following results were obtained:
metabolite |
amount |
specific radioactivity
(µCi /mmol) |
|||
µmol |
from [14C-U]glucose |
from [32P]ATP |
from [32P] inorganic phosphate |
||
glucose |
100 ± 5 |
0.99 ± 0.02 |
- |
- |
|
glucose 6-phosphate |
3.2 ± 0.5 |
0.98 ±
0.03 |
0.99 ±
0.03 |
0.98 ±
0.05 |
|
fructose 6-phosphate |
2.6 ± 0.6 |
0.99 ± 0.03 |
0.98 ± 0.03 |
0.99 ± 0.04 |
|
fructose 1,6-bisphosphate |
3.4 ± 0.7 |
0.99 ± 0.02 |
1.99 ± 0.04 |
1.98 ± 0.03 |
|
|
glyceraldehyde 3-phosphate |
2.9 ± 0.3 |
0.48 ± 0.03 |
0.98 ± 0.02 |
0.96 ± 0.02 |
|
dihydroxyacetone phosphate |
2.8 ± 0.2 |
0.47 ± 0.02 |
0.97 ± 0.03 |
0.96 ± 0.03 |
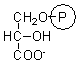 |
3-phosphoglycerate |
6.1 ± 0.3 |
0.49 ± 0.03 |
0.99 ± 0.02 |
0.96 ± 0.03 |
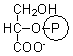 |
2-phosphoglycerate |
5.7 ± 0.2 |
0.51 ± 0.02 |
0.97 ± 0.03 |
0.98 ± 0.03 |
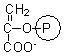 |
phosphoenolpyruvate |
6.2 ± 0.3 |
0.49 ± 0.03 |
0.99 ± 0.02 |
0.98 ± 0.03 |
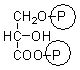 |
1,3-bisphosphoglycerate |
5.9 ± 0.2 |
0.47 ± 0.03 |
0.98 ± 0.01 |
1.99 ± 0.04 |
pyruvate |
6.4 ± 0.3 |
0.49 ± 0.01 |
- |
- |
|
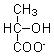 |
lactate |
160 ± 4 |
0.50 ± 0.02 |
- |
- |
ATP |
219 ± 3 |
- |
0 |
1.01 ± 0.02 |
|
ADP |
0 |
- |
- |
- |
|
The compounds shown in red are new intermediates that were not detected previously; they are shown in the order in which they elute from the high pressure liquid chromatography column.
The compounds shown in blue are those for which we are reasonably certain (from previous experiments) that this is the sequence in which they are formed.
What conclusions can you draw from these results?
As expected, under these conditions all of the ADP has been phosphorylated to ATP, and 2 mol of ATP have been formed for each mol of glucose consumed. Most of the glucose has been metabolised to lactate.
When [32P]ATP is used, both phosphate groups in fructose 1,6-bisphosphate arise from ATP, and this leads to one labelled phosphate in each of the phosphorylated intermediates. This suggests that the second phosphate incorporated into 1,2-bisphoglycerate has come from inorganic phosphate, not ATP.
This is confirmed by the experiments using [32P]inorganic phosphate - the specific activity of phosphate in 1,3-bisphosphoglycerate is double that of the other phosphorylated intermediates.
If the phosphate in fructose 1,6-biphosphate has come from ATP, as suggested by the experiments using [32P]ATP, why is it labelled in the experiments using [32P]inorganic phosphate?
There is net phosphorylation of ADP to ATP, and the experiments with [32P]inorganic phosphate show that the ATP that is formed incorporated the label from inorganic phosphate. This means that after a few reaction cycles most or all of the ATP present will be labelled with 32P.
 We have
seen previously that fructose 1,6-bisphosphate is cleaved to yield dihydroxyacetone
phosphate and glyceraldehyde 3-phosphate, which accumulate in equal amounts
when there is no NAD in the incubation medium. However, when NAD is added, neither
accumulates to any significant extent.
We have
seen previously that fructose 1,6-bisphosphate is cleaved to yield dihydroxyacetone
phosphate and glyceraldehyde 3-phosphate, which accumulate in equal amounts
when there is no NAD in the incubation medium. However, when NAD is added, neither
accumulates to any significant extent.
How can you begin to deduce the sequence of reactions involved in the metabolism of dihydroxyacetone phosphate and glyceraldehyde 3-phosphate?
One obvious experiment would be to add either dihydroxyacetone phosphate or glyceraldehyde 3-phosphate to the incubations with and without added NAD.
The following results were obtained:
No added NAD |
+ 20 µmol NAD |
|||
| + 20 µmol dihydroxyacetone phosphate | + 20 µmol glyceraldehyde 3-phosphate |
+ 20 µmol dihydroxyacetone phosphate | + 20 µmol glyceraldehyde 3-phosphate |
|
| dihydroxyacetone phosphate | 9.8 ± 0.2 µmol |
9.9 ± 0.2 µmol |
0.5 ± 0.05 µmol |
0.1 ± 0.01 µmol |
| glyceraldehyde 3-phosphate | 9.9 ± 0.3 µmol |
9.8 ± 0.2 µmol |
3.9 ± 0.2 µmol |
3.8 ± 0.3 µmol |