Metabolism
on-line - the virtual tutorial room
copyright © 2008 - 2015 David A Bender
An adverse response to antimalarial medication -
 Samuel
W is an African-American recruit to the army. He was given the antimalarial
drug primaquine, and suffered a delayed reaction with kidney pain, dark urine,
and low red blood cell counts that led to anaemia and weakness. Centrifugation
of a blood sample showed a low haematocrit, and the plasma was red coloured.
Samuel
W is an African-American recruit to the army. He was given the antimalarial
drug primaquine, and suffered a delayed reaction with kidney pain, dark urine,
and low red blood cell counts that led to anaemia and weakness. Centrifugation
of a blood sample showed a low haematocrit, and the plasma was red coloured.
Similar acute haemolytic attacks have been observed, predominantly in men of Afro-Caribbean origin, in response to primaquine and a variety of other drugs, including dapsone, the antipyretic acetylphenylhydrazine, the antibacterial bactrim/septrim, sulphonamides and sulphones, whose only common feature is that they all undergo cyclic non-enzymic reactions in the presence of oxygen to produce hydrogen peroxide and a variety of oxygen radicals that can cause oxidative damage to membrane lipids, leading to haemolysis. Moderately severe infection can also precipitate a haemolytic crisis in susceptible people.
Why do you think that infection can lead to a haemolytic crisis in susceptible people?
Part of the response to infection is activation of macrophages to produce a mixture of oxygen and other radicals that are cytotoxic to engulphed micro-organisms. This leads to an increase in whole body radical burden, and increased oxidative damage to cell membranes.
One way of screening for sensitivity to primaquine is based on the observation that the glutathione concentration of erythrocytes from sensitive subjects is somewhat lower than that of control subjects, and falls considerably on incubation with acetylphenylhydrazine.
Glutathione (GSH) is a tripeptide, gamma-glutamyl-cysteinyl-glycine, which readily undergoes oxidation to form a disulphide-linked hexapeptide, oxidised glutathione, generally abbreviated to GSSG

The table below shows the concentrations of GSH and GSSG in red cells from 10 control subjects, and Samuel W, before and after incubation with acetylphenylhydrazine.
The effect of incubation with 330 µmol /L acetylphenylhydrazine on erythrocyte glutathione.
controls |
Samuel W |
|||
GSH mmol /L |
GSSG µmol /L |
GSH mmol /L |
GSSG µmol /L |
|
| initial | 2.01 ± 0.29 |
4.2 ± 0.61 |
1.61 |
400 |
| + acetylphenylhydrazine | 1.82 ± 0.24 |
190 ± 28 |
0.28 |
1540 |
What conclusions can you draw from these results?
How much glutathione has been oxidised per mol of acetylphenylhydrazine added?
Samuel’s red blood cells contain significantly less GSH than normal, and a very high level of GSSG.
The results suggest that acetylphenylhydrazine causes the oxidation of glutathione to GSSG. This is not a simple stoichiometric oxidation of glutathione by acetylphenylhydrazine. In Samuel's erythrocytes, the ratio of glutathione oxidised : acetylphenylhydrazine present = 4.7. This suggests that it is likely that acetylphenylhydrazine undergoes a cyclic redox reaction that results in the production of hydrogen peroxide, which in turn oxidises glutathione.
In control subjects incubation with acetylphenylhydrazine leads to a modest decrease in GSH, and a small accumulation of GSSG; Samuel’s red cells show a very considerable depletion of GSH and a very large accumulation of GSSG on incubation with acetylphenylhydrazine.
What might explain this very large accumulation of GSSG in the presence of acetylphenylhydrazine?
This suggests that Samuel cannot reduce GSSG back to GSH as effectively as normal. This could be due to lack of either glutathione reductase or NADPH.
The reported Km of glutathione reductase for GSSG is 65 µmol /L and for NADPH 8.5 µmol /L Erythrocyte lysates were incubated with a saturating concentration of GSSG (1 mmol /L) and either NADPH or NADH (100 µmol /L). Each incubation contained the haemolysate from 0.5 mL packed cells.
Glutathione reductase, µmol product formed /min
controls |
Samuel W |
|
| NADPH | 0.63 ± 0.06 |
0.64 |
| NADH | 0.01 ± 0.001 |
0.01 |
What conclusions can you draw from these results?
Obviously Samuel has no problem with the activity of glutathione reductase, suggesting that his problem lies in providing a source of NADPH for the enzyme.
Since none of the lysates showed any significant activity with NADH, it is unlikely that there is any transhydrogenase activity in erythrocytes, to reduce NADP+ to NADPH at the expense of NADH. This raises the problem of the source of NADPH in erythrocytes.
The dye methylene blue will oxidise NADPH; the reduced dye then undergoes non-enzymic oxidation in air, so the addition of a relatively small amount of methylene blue will effectively deplete NADPH, and would be expected to stimulate any pathway that reduces NADP+ to NADPH.
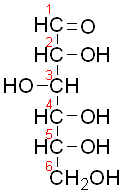 Erythrocytes
from control subjects were incubated with 10 mmol /L [14C]glucose with or without
the addition of methylene blue; all 6 possible positional isomers of [14C]glucose
were used. The radioactivity in (lactate plus pyruvate) was determined after
thin layer chromatography of the incubation mixture, and radioactivity in any
carbon dioxide formed was also measured. Each incubation contained 1 mL erythrocytes
in a total incubation volume of 2 mL.
Erythrocytes
from control subjects were incubated with 10 mmol /L [14C]glucose with or without
the addition of methylene blue; all 6 possible positional isomers of [14C]glucose
were used. The radioactivity in (lactate plus pyruvate) was determined after
thin layer chromatography of the incubation mixture, and radioactivity in any
carbon dioxide formed was also measured. Each incubation contained 1 mL erythrocytes
in a total incubation volume of 2 mL.
Production of [14C] (lactate +pyruvate) and carbon dioxide by 1 mL erythrocytes incubated for 1 hour with 10 mmol /L [14C]glucose at 10 µCi /mmol. Figures show mean dpm (radioactive disintegrations per minute) ± sd for 5 replicate incubations.
control |
+ methylene blue |
|||
lactate + pyruvate |
carbon dioxide |
lactate + pyruvate |
carbon dioxide |
|
| [14C-1]glucose | 12680 ± 110 |
1410 ± 15 |
1830 ± 20 |
12260 ± 130 |
| [14C-2]glucose | 14080 ± 120 |
ND |
14120 ± 120 |
ND |
| [14C-3]glucose | 14100 ± 120 |
ND |
14090 ± 120 |
ND |
| [14C-4]glucose | 14060 ± 120 |
ND |
14080 ± 120 |
ND |
| [14C-5]glucose | 14120 ± 120 |
ND |
14060 ± 120 |
ND |
| [14C-6]glucose | 14190 ± 110 |
ND |
14100 ± 120 |
ND |
ND = not detectable – i.e. below the limits of detection.
What conclusions can you draw from these results?
Does methylene blue affect the total amount of glucose that has been metabolised?
Considering the control incubations: only carbon-1 of glucose gives rise to carbon dioxide, and the radioactivity in (lactate + pyruvate) is lower from [14C-1]glucose than the other positional isomers. However, the total radioactivity in (lactate + pyruvate + carbon dioxide) is ~14090 dpm in all cases - from the initial specific activity this gives a rate of glucose utilisation of 640 nmol /mL cells /hour.
There is no reaction in glycolysis in which carbon dioxide is produced, and this suggests that there must be some other pathway of glucose metabolism in the erythrocyte, in which carbon-1 of glucose is released as carbon dioxide. In the control incubations, about 10% of the radioactivity from carbon-1 of glucose appears as carbon dioxide, suggesting that perhaps 10% of glucose metabolism is by way of this alternative pathway.
In the presence of methylene blue there is a very considerable increase in the production of [14C]carbon dioxide from [14C-1]glucose, and a corresponding decrease in the label in lactate and pyruvate, although the total radioactivity in (lactate + pyruvate + carbon dioxide) is the same as in the control incubation. This suggests that the overall rate of glucose metabolism is unaffected by methylene blue, but a greater proportion (87%) is by way of the decarboxylation pathway.
Since the action of methylene blue is to oxidise NADPH, it is therefore likely that the pathway that results in decarboxylation is also the pathway that utilises NADP+ and forms NADPH.
Two reactions involving glucose metabolites catalyse the reduction of NADP+ to NADPH: glucose 6-phosphate dehydrogenase, forming 6-phosphogluconate, and phosphogluconate dehydrogenase, which yields the 5-carbon sugar ribulose 5-phosphate, liberating carbon dioxide from carbon-1 of the substrate.

These reactions are the first two reactions in the pentose phosphate pathway, which provides an alternative to part of the pathway of glycolysis, yielding glyceraldehyde 3-phosphate and fructose 6-phosphate. This pathway is a major source of NADPH, and in addition to its importance in red blood cells, it provides about half the NADPH required for fatty acid synthesis. It is also the pathway for synthesis of the 5-carbon sugar ribose. Click here to see the complete pathway.