Metabolism
on-line - the virtual tutorial room
copyright © 2008 - 2015 David A Bender
Breathless after sprinting
WS is a sprinter; he weighs 75 kg. At the end of a 100 m race he is breathing rapidly and deeply, and continues to do so for several minutes. His plasma lactate and pyruvate were measured before and immediately after the race, and again 30 min later, when his breathing had returned to normal.
lactate (mmol /L) |
pyruvate
(mmol /L) |
|
| before the race | 0.5 |
0.11 |
| immediately after | 11.5 |
0.09 |
| 30 minutes later | 1.0 |
0.12 |
Can you explain why he was hyperventilating at the end of the race?
For reasons that we will be exploring in the exercise, he is acidotic, with a high plasma concentration of lactate. Note, however, that unlike the cases of PC and BD, who were considered in the exercise on Life-threatening acidosis in an alcoholic - and in a hunger striker given intravenous glucose, WS's plasma pyruvate is within the normal range at all times.
The usual response to acidosis is to increase the rate of breathing, so as to expel carbon dioxide, and so shift the equilibrium below to the left, lowering the hydrogen ion concentration in the bloodstream, and so raising plasma pH.
![]()
His oxygen consumption was also measured for 10 minutes before the race, and over the 30 minutes after the race. At rest he was consuming 0.5 L oxygen per minute. Over the 30 minutes after the race his average oxygen consumption was 14% higher, with a total consumption over 30 minutes of 17.12 litres of oxygen.
What we have to consider is where the lactate has come from, how it disappears during the 30 minutes after the race and why he consumes 14% more oxygen over the thirty minutes after he has completed the race.
In a series of experiments with anaesthetised fasting dogs cannulae were placed in the femoral artery and popliteal vein to permit measurement of metabolites taken up, or put out, by the gastrocnemius-plantaris muscle group after gentle electrical stimulation (1 twitch per second) and after vigorous electrical stimulation (5 twitches per second). Cannulae were also placed in the hepatic artery and hepatic vein, to permit measurement of metabolites taken up and put out by the liver. The result are shown as the arterio-venous difference as nmol /g tissue /minute; positive values indicate output from the tissue, negative values indicate uptake by the tissue. Figures show mean ± sd for 3 x replicate experiments.
muscle |
liver |
|||
gentle stimulation |
vigorous stimulation |
gentle stimulation |
vigorous stimulation |
|
| glucose | -215 ± 12 |
-885 ± 15 |
+100 ± 12 |
+660 ± 20 |
| oxygen | -4515 ± 50 |
-6912 ± 50 |
-1150 ± 50 |
-1800 ± 50 |
| lactate | +20 ± 5 |
+1112 ± 50 |
-50 ± 10 |
-1100 ± 40 |
What conclusions can you draw from these results?
In response to gentle stimulation muscle is mainly aerobic - less than 10% of the glucose taken up is released as lactate, with the remainder being metabolised to carbon dioxide and water.
In response vigorous stimulation there is the expected large increase in uptake of both glucose and oxygen, but there is also a large increase in lactate output, so that now more than 60% of the glucose taken up is released as lactate.
When muscle is only gently stimulated, the liver is putting out a small amount of glucose - presumably this is arising partly from its glycogen reserves, and partly as a result of gluconeogenesis from various substrates.
When muscle is stimulated vigorously, the liver takes up a large amount of lactate, and puts out a large amount of glucose. Oxygen consumption also increases at this time.
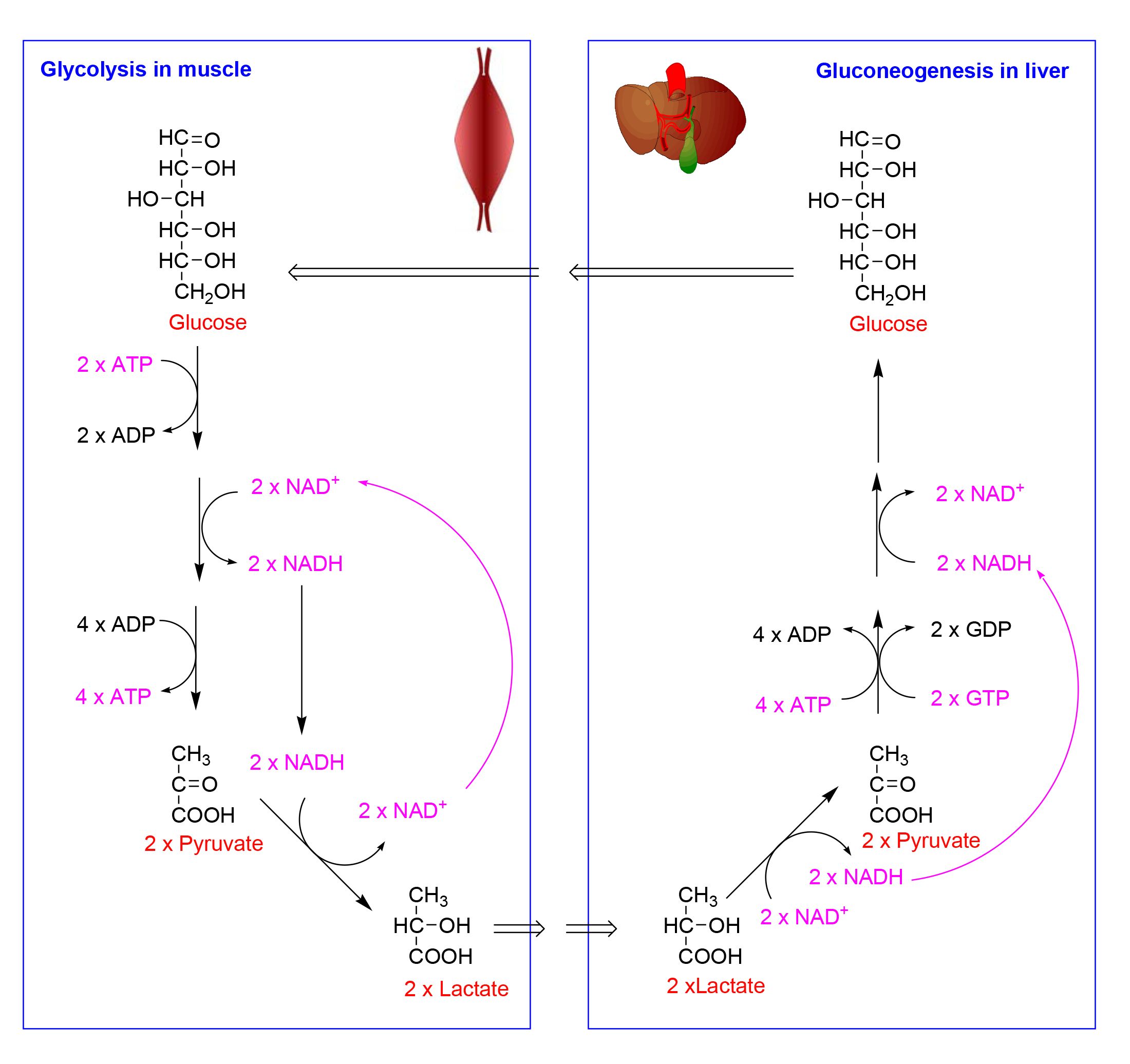
We can therefore conclude that in vigorous exercise muscle metabolises as much glucose aerobically as it can, but is presumably limited by the availability of oxygen, and in order to produce enough ATP to maintain a high rate of work output, it metabolises much glucose anaerobically, forming lactate in order to re-oxidise the NADH formed in glycolysis.
We can also conclude that the lactate is taken up by the liver, and is used for resynthesis of glucose, which is then exported, and presumably taken up by the muscle, either for immediate use or for synthesis of glycogen reserves in the muscle.
We can summarise this as an inter-organ metabolic cycle of anaerobic glycolysis in the muscle and gluconeogenesis in the liver.
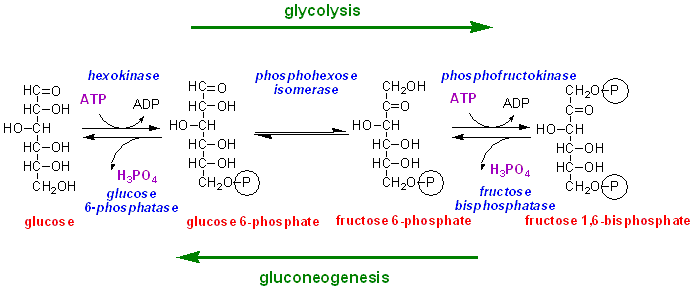
We now need to consider what is involved in the conversion of lactate back to glucose.
There are three steps in glycolysis that are irreversible under physiological conditions:
The phosphorylation of glucose to glucose 6-phosphate, catalysed by hexokinase (and in the liver also by glucokinase)
The phosphorylation of fructose 6-phosphate to fructose 1,6-bisphosphate catalysed by phosphofructokinase
The reaction catalysed by pyruvate kinase, in which ADP is phosphorylated to ATP by transfer of phosphate from phosphoenolpyruvate.
The first two reactions are irreversible under physiological conditions because the ratio of ATP : ADP is ~ 500 : 1, and ATP binds very much better to these enzymes than does ADP.
For the first two reactions, there are separate enzymes involved in gluconeogenesis, glucose 6-phosphatase and fructose 1,6-bisphosphatase, which catalyse simple hydrolysis of the phosphate group, releasing inorganic phosphate. Fructose 1,6-bisphosphatase and glucose 6-phosphatase are only found in tissues that catalyse gluconeogenesis (liver, kidney and possibly the small intestine)